Zagadnienia
11. Modele z zależnością przestrzenną: dyfuzja w procesach biologicznych
Do tej pory rozważaliśmy modele populacyjne, w których nie uwzględnialiśmy zależności od położenia osobników w przestrzeni, a tylko średnie zagęszczenia i ich zmiany w czasie. Takie podejście do modelowania dynamiki populacji kryje za sobą dodatkowe założenia dotyczące równomiernego rozłożenia przestrzennego osobników danej populacji oraz braku istotnego wpływu zmian położenia na opisywany proces. Oczywiste wydaje się, że w wielu przypadkach położenie i ruch w przestrzeni odgrywają dużą, jeśli nie zasadniczą rolę w przebiegu danego procesu. W związku z tym kolejną część wykładu poświęcimy omówieniu jednego z możliwych sposobów modelowania ruchu, tzw. procesu dyfuzji. W przypadku procesów chemicznych dyfuzja opisuje ruch substancji w kierunku od większych do mniejszych stężeń. Możliwe są oczywiście także przepływy innego typu — np. chemotaksja działa w pewnym sensie przeciwnie do dyfuzji.
Wyprowadzimy najpierw równanie dyfuzji substancji chemicznej w przestrzeni jednowymiarowej (dla przestrzeni dwu i trójwymiarowej rozumowanie przebiega analogicznie). Załóżmy, że mamy bardzo wąską rurkę o długości
Z punktem
w punkcie
w punkcie
Stąd, dzieląc stronami powyższą równość przez
| (11.1) |
czyli jednowymiarowe równanie dyfuzji. W ogólnym przypadku równanie dyfuzji ma postać
gdzie
Z naszego populacyjnego punktu widzenia będziemy rozpatrywać dyfuzję jako proces losowego przemieszczania się osobników w przestrzeni. Najprostsze podejście do tego typu zagadnień reprezentowane jest przez tzw. ruchy Browna. Ponownie dla uproszczenia omówimy sytuację jednowymiarową. Rozważamy prostą podzieloną na odcinki o długości
| (11.2) |
gdzie
Wstawiając powyższe rozwinięcia do równania (11.2) dostajemy
Załóżmy teraz, że
Dzieląc to równanie przez
Otrzymaliśmy więc związek losowego przemieszczenia się osobników z równaniem dyfuzji z jednej strony i pewnym łańcuchem Markowa z drugiej.
11.1. Równania ewolucyjne: równanie Fishera – Kołmogorowa
W przypadku rzeczywistych populacji oprócz przemieszczania się rozpatrujemy zwykle także inne procesy, np. rozrodczości/śmiertelności. W ogólnym przypadku otrzymujemy równanie reakcji-dyfuzji
| (11.3) |
zwane także równaniem ewolucyjnym, które opisuje zmiany zagęszczenia populacji w chwili
Równanie Skellama
Najprostsze tego typu równanie z liniową kinetyką zostało użyte przez Skellama w 1951 roku do opisu rozprzestrzeniania się populacji. Dokładniej, za pomocą równania (11.3) z funkcją
Rozpatrzmy równanie dyfuzji (11.1) w całym
którą nazywamy rozwiązaniem fundamentalnym. Oczywiście spełnienie warunku początkowego należy rozumieć w sensie granicznym.
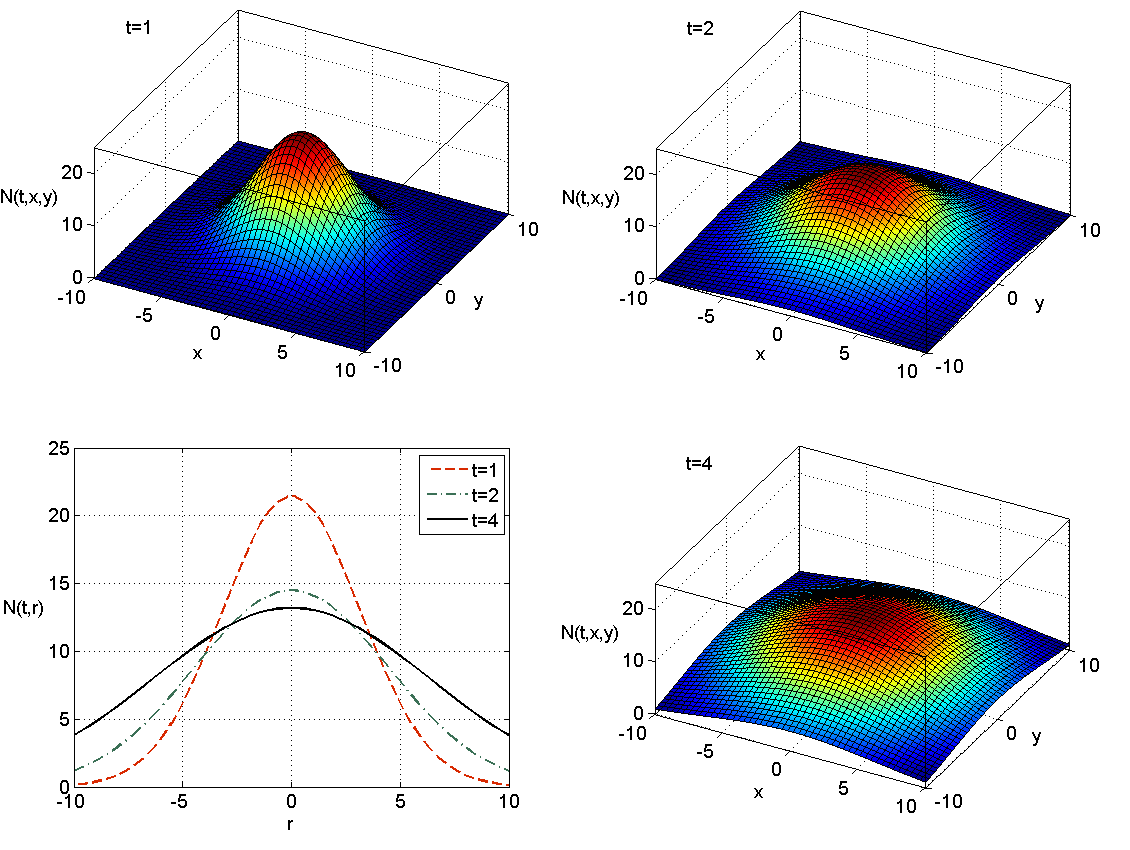
Ogólniej, jeśli rozważamy dyfuzję w przestrzeni
Teraz połączymy znajomość rozwiązania fundamentalnego równania dyfuzji dla
które zadaje falę inwazyjną. W punktach o ustalonym promieniu
Równanie Fishera – Kołmogorowa
Równanie Fishera [3] lub inaczej Fishera – Kołmogorowa jest z kolei najprostszym nieliniowym równaniem typu ewolucyjnego i stanowi naturalne uogólnienie równania logistycznego na przypadek populacji złożonej z osobników, które mogą się przemieszczać i ruch ten ma znaczenie dla opisywanego procesu. Jest to także prototypowe równanie, w którym występują rozwiązania w postaci fal biegnących. Typowo jest to równanie rozpatrywane w całej przestrzeni
Równanie to ma postać
| (11.4) |
przy czym w najprostszym przypadku, który omówimy,
Pokażemy, że równanie (11.4) ma rozwiązania w postaci fal biegnących. Zaczniemy od przeprowadzenia ubezwymiarowienia w celu zredukowania liczby parametrów. Przeprowadzając zamianę zmiennych
i wracając do standardowych oznaczeń zmiennych dostajemy
| (11.5) |
Odpowiadające równaniu (11.5) równanie kinetyczne, czyli równanie logistyczne, ma dwa stany stacjonarne, niestabilny
Definicja 11.1
Falą biegnącą nazywamy nietrywialne, ograniczone rozwiązanie równania (11.3) postaci
dla pewnego
Zauważmy, że
Uwaga 11.1
Równanie (11.1) nie ma rozwiązań w postaci fali biegnącej.
Załóżmy, że takie rozwiązanie
zatem wartości własne tego równania spełniają równanie kwadratowe
Dla równania Fishera – Kołmogorowa (11.5) rozwiązanie w postaci fali biegnącej musi spełniać równanie
| (11.6) |
i postulujemy, że
Równanie (11.6) jest równoważne układowi równań
| (11.7) |
gdzie prim (
Badając lokalną stabilność wyznaczamy wielomian charakterystyczny
i wartości własne
skąd wynika, że
Stwierdzenie 11.1
Dla
Dla układu (11.7) przy
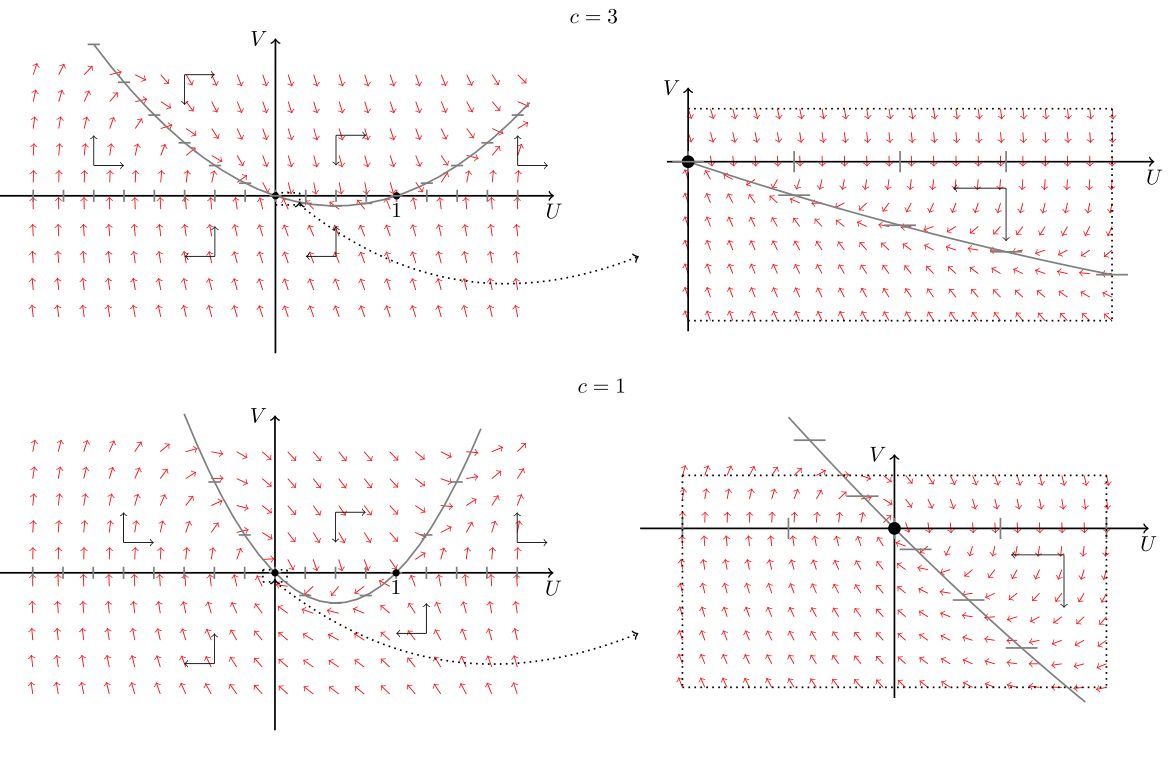
Uwaga 11.2
Dla
Zagadnienie występowania fal biegnących (bądź też fal innego typu) jest często badane w kontekście tego typu równań. Należy jednak zdawać sobie sprawę z tego, że nie zawsze rozważana dziedzina jest na tyle duża, że można założyć iż jest to cała przestrzeń. Wtedy, przy założeniu ograniczonej dziedziny, rozważamy zagadnienia początkowo-brzegowe.
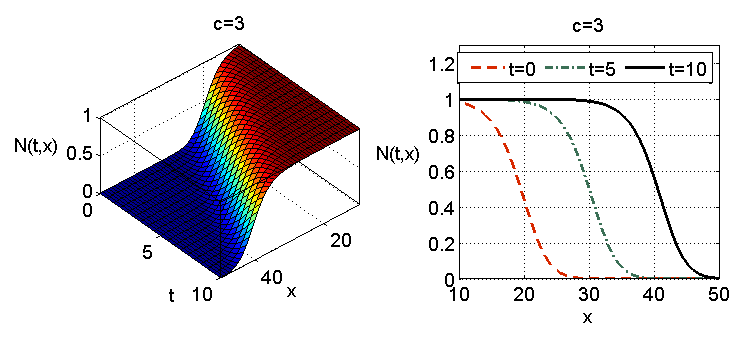
Równanie (11.4) było także rozpatrywane w kontekście rozwoju nowotworu przy założeniu warunku brzegowego. Załóżmy dla uproszczenia jednowymiarową propagację komórek na odcinku
| (11.8) |
i warunkiem brzegowym Neumanna, czyli
| (11.9) |
Dla zagadnienia początkowo-brzegowego (11.4) (11.8) (11.9) lokalne istnienie i jednoznaczność wynika z własności kinetyki.
Korzystając z twierdzenia porównawczego 11.1 i własności rozwiązań równania logistycznego możemy wykazać, że dla warunku początkowego
Twierdzenie 11.1
Niech
gdzie
-
jeśli
-
jeśli
Wtedy albo
Badając dynamikę modelu zwykle zaczynamy od zbadania stabilności rozwiązań stacjonarnych. Dla modeli niejednorodnych przestrzennie, oprócz rozważanych do tej pory rozwiązań stacjonarnych niezależnych od przestrzeni możemy mieć także rozwiązania stacjonarne, które zależą od położenia w dziedzinie równania. Jednak typowo analizę takiego zagadnienia zaczynamy od zbadania istnienia i stabilności rozwiązań stacjonarnych jednorodnych przestrzennie, a potem przechodzimy do znacznie trudniejszych zagadnień związanych z niejednorodnością przestrzenną. Jednym z takich zagadnień jest powstawanie tzw. wzorów Turinga — od strony analitycznej wiąże się to z bifurkacją stabilnego przestrzennie stanu stacjonarnego do niestabilnego, kiedy w otoczeniu stanu stacjonarnego powstają skupiska osobników — grupy o różnym zagęszczeniu — tworzące wzory przestrzenne.
Niech
| (11.10) |
gdzie
W przypadku równania (11.4) możemy udowodnić znacznie więcej, czyli globalną stabilność
Twierdzenie 11.2
Jeśli
Twierdzenie porównawcze pokazuje, że
| (11.11) |
oraz
Wtedy
Mnożąc równanie (11.11) przez
Widzimy, że jeśli
11.2. Wzory Turinga
Jak już wspomnieliśmy, w przypadku jednego równania kinetycznego formowanie się wzorów Turinga nie jest możliwe, gdyż stabilność nie zmienia się pod wpływem dyfuzji. Natomiast dla
Badając możliwość występowania wzorów Turinga przeanalizujemy lokalną stabilność jednorodnych rozwiązań stacjonarnych w układzie dwóch równań reakcji-dyfuzji
| (11.12) |
gdzie
gdzie
Dla uproszczenia układ (11.12) będziemy rozpatrywać z jednorodnym warunkiem brzegowym Neumanna na
Badając stabilność stanu stacjonarnego zlinearyzujemy układ (11.12)
| (11.13) |
przy czym
Okazuje się, że liniowe równanie (11.13) można rozwiązać metodą rozdzielenia zmiennych, czyli
| (11.14) |
Podstawiając postać rozwiązania (11.14) do układu (11.13) otrzymujemy
| (11.15) |
skąd dostajemy układ równań na współczynniki
| (11.16) |
Rozwiązanie dostajemy przyrównując wyznacznik układu (11.16) do 0
| (11.17) |
W przypadku bez dyfuzji równanie (11.17) ma postać
i stabilność stanu stacjonarnego zależy od śladu i wyznacznika macierzy Jacobiego układu kinetycznego. Precyzyjniej, jeśli
-
[I]
-
-
to stan stacjonarny dla
dostajemy nierówność na
W analogiczny sposób możemy badać tworzenie się wzorów przestrzennych w modelu melanogenezy, przy czym wielkość i różnorodność formujących się wzorów zależy oczywiście od parametrów modelu. Jeśli dla danego gatunku dominują składniki kinetyczne, to powstające wzory są powtarzalne (jak np. u żyraf czy zebr), a jeśli składniki losowe, to wzory mogą się bardzo różnić w zależności od osobników (jak w przypadku kotów domowych).



