Zagadnienia
12. Modelowanie odpowiedzi odpornościowej
Dwa kolejne wykłady zostaną poświęcone modelowaniu reakcji odpornościowych. Na początku przedstawimy podstawowe informacje dotyczące działania układu odpornościowego człowieka.
12.1. Układ odpornościowy człowieka
Układ odpornościowy człowieka (i nie tylko człowieka) jest bardzo skomplikowany. Odpowiedź odpornościową wywołuje antygen. Antygenem nazywamy substancję, która wykazuje się:
-
immunogennością, czyli zdolnością indukowania swoistej (specyficznej) odpowiedzi odpornościowej przeciw sobie;
-
antygenowością, czyli zdolnością do swoistego łączenia się z immunoglobulinami (zarówno wolnymi jak i stanowiącymi receptory limfocytów $B$) oraz receptorami limfocytów $T$.
Okazuje się, że w obrębie jednego antygenu może znajdować się wiele miejsc wiązanych przez przeciwciała. Są to tzw. epitopy lub determinanty antygenowe. Epitopy obecne w jednej cząsteczce antygenu mogą być identyczne lub różne i mogą być wiązane przez przeciwciała o tej samej lub różnej swoistości. Antygen zawierający wiele epitopów nazywamy wielowartościowym lub poliwalentnym.
Reakcja odpornościowa składa się z wielu różnorodnych procesów, w których bierze udział wiele składników układu odpornościowego. Najważniejsze z tych składników to:
-
wśród komórek odpornościowych:
-
limfocyty
-
limfocyty
-
komórki prezentujące antygen, ang. antigen presenting cells (APC);
-
wśród cząstek:
-
immunoglobuliny (przeciwciała);
-
receptory limfocytów wiążące się z antygenem.
Zestawienie najważniejszych składników układu odpornościowego przedstawiamy na rys. 12.1 i w tabeli 12.1.
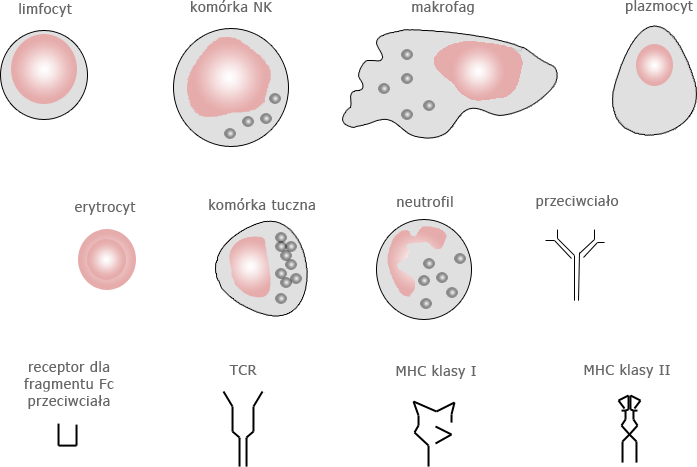
| Limfocyty |
rozpoznają antygeny i wytwarzają przeciwciała |
|---|---|
| Komórki plazmatyczne (plazmocyty) | wytwarzają przeciwciała |
| Limfocyty |
wspomagają odpowiedź odpornościową; wydzielają cytokiny |
| Limfocyty |
hamują odpowiedź odpornościową; wydzielają cytokiny |
| Limfocyty |
są zdolne do zabijania komórek docelowych |
| Komórki |
są zdolne do cytotoksyczności komórkowej zależnej od przeciwciał |
| Komórki |
są zdolne do spontanicznego zabijania komórek nowotworowych i zakażonych przez wirusy |
| Komórki dendrytyczne | prezentują antygeny limfocytom |
| Monocyty, makrofagi | są zdolne do fagocytozy i zabicia mikroorganizmów oraz innych obcych komórek; wydzielają cytokiny |
| Neutrofile | są zdolne do fagocytozy i zabijania mikroorganizmów |
| Komórki tuczne, bazofile | biorą udział w nadwrażliwości I (anafilaksji) |
| Eozynofile | są zdolne do fagocytozy i zbijania niektórych pasożytów z użyciem przeciwciał |
| Megakariocyty | wytwarzają trombocyty, które uczestniczą w krzepnięciu krwi i reakcjach zapalnych |
| Erytrocyty | poprzez receptor dla dopełniacza wiążą kompleksy immunologiczne i uczestniczą w ich usuwaniu |
W każdej odpowiedzi odpornościowej oprócz komórek efektorowych (np. limfocytów
Rodzinę komórek zwanych limfocytami
-
limfocyty $T$ cytotoksyczne (
-
limfocyty $T$ pomocnicze, ang.
-
limfocyty $T$ regulatorowe, ang. regulatory
Z kolei limfocyty
Typy odpowiedzi odpornościowej
Odpowiedź odpornościową możemy podzielić na:
-
swoistą (specyficzną) i nieswoistą (niespecyficzną);
-
typu humoralnego i komórkowego.
Mechanizmy nieswoiste nazywane są też odpornością natywną lub wrodzoną. Jednak żadna z tych nazw nie oddaje istoty tego typu reakcji. Podobnie zresztą dzieje się w przypadku mechanizmów swoistych nazywanych też odpornością adoptywną.
Mechanizmy nieswoiste rozwinęły się wcześniej w filogenezie (tzn. są starsze ewolucyjnie), są mniej precyzyjne, ale reakcje zachodzą szybko. Mówi się, że stanowią pierwszą linię obrony organizmu. Z kolei mechanizmy swoiste są filogenetycznie młodsze. W trakcie reakcji odpornościowej pojawienie się tych mechanizmów wymaga czasu, ale nakierowane są dokładnie przeciw konkretnemu rodzajowi antygenu, więc działają precyzyjniej i skuteczniej.
W odpowiedzi swoistej biorą udział głównie antygeny swoiste wyprodukowane przez komórki plazmatyczne powstałe w procesie rozpoznawania antygenu przez limfocyty
Z kolei podział na odpowiedź humoralną i komórkową jest cokolwiek sztuczny. W większości przypadków obserwuje się oba typy reakcji, chociaż w konkretnych przypadkach dla danego antygenu jedna z nich może dominować nad drugą.
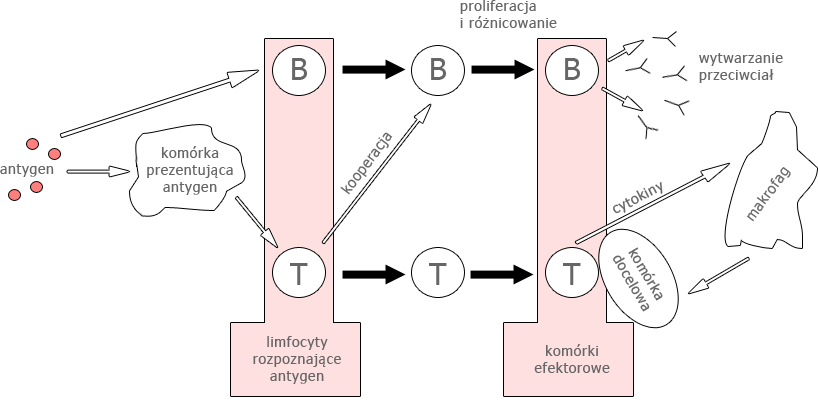
W odpowiedzi humoralnej decydującą rolę w zwalczaniu antygenu odgrywają przeciwciała obecne głównie we krwi odwodowej i limfie, ale także w innych płynach organicznych, natomiast za odpowiedź komórkową odpowiadają limfocyty
12.2. Proste modele odpowiedzi odpornościowej
Zajmiemy się teraz zaproponowaniem możliwie najprostszych modeli odpowiedzi odpornościowej, gdzie ponownie wykorzystamy znane już sposoby modelowania oddziaływań między różnymi populacjami. Zacznijmy od opisu wewnętrznej dynamiki antygenu. Pod względem tejże dynamiki możemy antygeny podzielić na:
-
aktywne, jak bakterie, grzyby i wirusy, mające zdolność aktywnego namnażania się w organizmie;
-
pasywne, jak trucizny i jady, które się nie namnażają;
-
własne, czyli produkowane przez organizm, jak autoantygeny czy antygeny transplantacyjne.
Niech
| (12.1) |
gdzie
| (12.2) |
gdzie
Zakładamy teraz, że dany antygen wywołuje odpowiedź odpornościową, którą w modelu reprezentują efektory (limfocyty lub przeciwciała) o zagęszczeniu
gdzie
Układ odpornościowy aktywuje się po wykryciu antygenu przez komórki odpornościowe. Wiemy też, że efektory zużywają się w trakcie odpowiedzi odpornościowej, zatem równanie ich dynamiki możemy zapisać w następującej postaci
gdzie
Ostatecznie model odpowiedzi odpornościowej przeciw antygenowi aktywnemu opiszemy następującym układem dwóch równań różniczkowych zwyczajnych
| (12.3) |
z dodatnimi współczynnikami. Analogicznie możemy modelować odpowiedź przeciwko antygenowi pasywnemu oraz autoantygenowi, co pozostawiamy jako ćwiczenie.
Przejdziemy teraz do analizy układu (12.3). Podstawowe własności jak istnienie, jednoznaczność, nieujemność i przedłużalność rozwiązań badamy w sposób standardowy. Istnienie i jednoznaczność rozwiązań dla dowolnego warunku początkowego
W kolejnym kroku analizy znajdziemy izokliny zerowe i naszkicujemy portrety fazowe w zależności od parametrów układu (12.3). Widzimy, że
Zauważmy, że oś
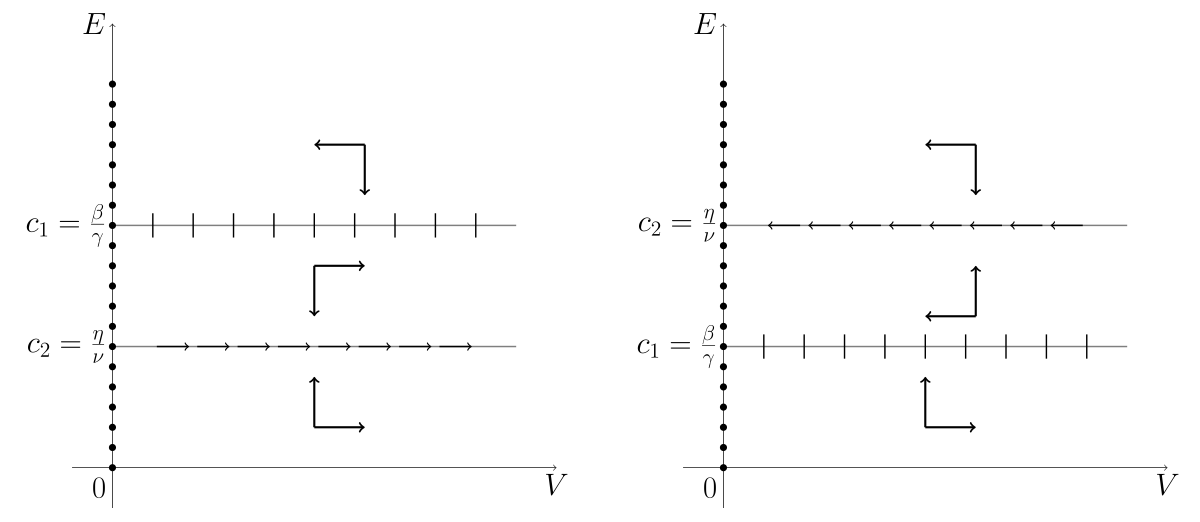
Oznaczmy
| (12.4) |
Zauważmy, że
Jeśli
skąd
| (12.5) |
Rozpatrzmy przypadek
| (12.6) |
Taka dynamika jest w tym przypadku możliwa pod warunkiem, że
| (12.7) |
Wniosek 12.1
Jeśli
Wniosek 12.1 pokazuje, że jeśli antygen jest silny (spełnia nierówność
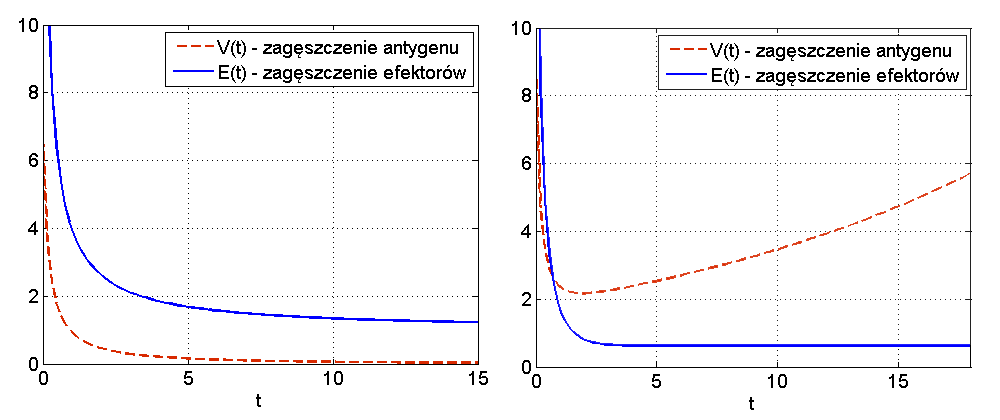
W przypadku słabego antygenu lub dostatecznie silnego układu odpornościowego, czyli dla
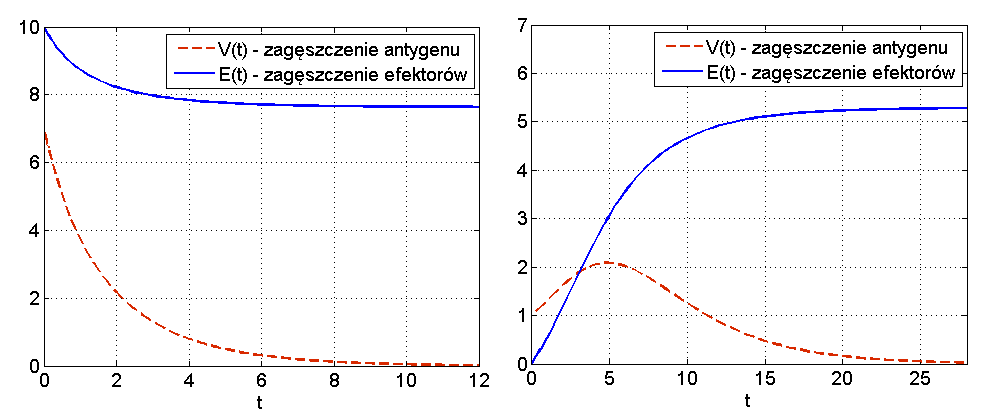
Widzimy więc, że układ (12.3) jest zbyt prosty, aby mógł w odpowiedni sposób odzwierciedlić rzeczywiste procesy. W szczególności w rzeczywistości nie jest możliwe, by nawet bardzo silny układ odpornościowy zawsze poradził sobie z antygenem, bez względu na wielkość dawki początkowej.


