13. Model Marczuka humoralnej odpowiedzi odpornościowej
Kolejny wykład poświęcimy na zapoznanie się z nieco bardziej skomplikowanym modelem odpowiedzi odpornościowej. W 1975 r. Guri I. Marczuk (właściwie G. I. Marchuk według obowiązującej transkrypcji rosyjskiego nazwiska, Marczuk to spolszczona wersja tego nazwiska) zaproponował dość prosty model, który jest w stanie odzwierciedlić najważniejsze przebiegi odpowiedzi odpornościowej, w tym również chorobę przewlekłą (chroniczną). Model opisany jest za pomocą czterech równań z opóźnionym argumentem i z założenia odzwierciedla humoralną odpowiedź odpornościową, choć podobną strukturę modelu można także zaproponować w przypadku odpowiedzi komórkowej. Model ten będziemy w dalszym ciągu nazywać modelem Marczuka. W 1980 r. ukazała się książka Marczuka [14], w której oprócz tego najprostszego modelu przedstawił także kilka wersji bardziej rozbudowanych, w tym model opisujący równoległy przebieg odpowiedzi humoralnej i komórkowej. W późniejszych latach ukazały się także tłumaczenia tej książki na język angielski [11] i polski [12]. Wyniki doświadczalne dla pewnych chorób okazały się bardzo podobne do teoretycznych wyników uzyskanych na podstawie modelu Marczuka.
Z powstaniem modelu wiąże się ciekawa historia. Marczuk przez wiele lat był przewodniczącym Akademii Nauk ZSSR. Niestety zachorował na żółtaczkę zakaźną, a pobyt w szpitalu urozmaicał sobie poznawaniem mechanizmów odpornościowych. W rezultacie doprowadziło to do powstania modelu reakcji odpornościowej i zaowocowało wieloma pracami naukowymi w tej dziedzinie. Po powrocie do zdrowia Marczuk w dalszym ciągu prowadził działalność naukową, w ramach której odwiedził Wydział Matematyki, Informatyki i Mechaniki UW, gdzie zaprezentował model i wyniki symulacji wskazujące możliwości leczenia choroby przewlekłej za pomocą zaostrzenia infekcji. Zarówno sam model, jak i możliwości jego zastosowania, zafascynowały jednego z pionierów polskiej biomatematyki, prof. Wiesława Szlenka. Tematyka ta znalazła się później wśród zagadnień badawczych w powstałym po pewnym czasie Instytucie Matematyki Stosowanej i Mechaniki.
13.1. Prezentacja modelu Marczuka
Zajmiemy się teraz omówieniem konstrukcji prostego modelu odpowiedzi odpornościowej zaproponowanego przez G. I. Marczuka, a następnie przedstawimy podstawowe własności układu.
Przypomnijmy, że reakcja odpornościowa następuje w wyniku wniknięcia do organizmu obcego czynnika, który nazywamy antygenem. W przypadku omawianego modelu antygenem są głównie bakterie, ponieważ np. wirusy wywołują znacznie bardziej skomplikowane reakcje, które wymagają bardziej skomplikowanego opisu matematycznego, w szczególności uwzględnienia także komórkowej odpowiedzi odpornościowej. Pojawienie się antygenu wywołuje kaskadę reakcji, które w efekcie doprowadzają do rozpoznania antygenu i uruchomienia produkcji przeciwciał swoistych przez komórki plazmatyczne. Mechanizmem uruchamiającym tę kaskadę jest powstanie tzw. kompleksu VT, czyli przyłączenie do antygenu przeciwciała prezentującego. Wobec tego zakładamy, że produkcja komórek plazmatycznych zależy od liczby wiązań antygen – przeciwciało, przy czym obserwujemy opóźnienie między powstaniem takiego wiązania a wyprodukowaniem przed układ odpornościowy komórek plazmatycznych produkujących przeciwciała swoiste.
W modelu występują następujące podstawowe zmienne
przy czym (jak dotychczas) używamy określenia zagęszczenie w stosunku do osobników pewnej populacji (tu populacji komórkowych), natomiast stężenie dotyczy głównie substancji, a wiemy, że przeciwciała są białkami.
Dynamikę antygenu opisujemy w modelu Marczuka w taki sam sposób jak w najprostszym modelu odpowiedzi odpornościowej prezentowanym poprzednio. Zakładamy, że bakterie rozmnażają się za stałym współczynnikiem reprodukcji
Przeciwciała giną nie tylko w wyniku wiązania z antygenem, ale też w procesie naturalnej degradacji. Są one produkowane przez komórki plazmatyczne w stałym tempie
gdzie
Zauważyliśmy wcześniej, że produkcja komórek plazmatycznych uruchamiana jest przez powstanie kompleksów
gdzie
Jeśli antygen występuje w organizmie, to w powyższym równaniu uwzględniamy jeszcze składnik związany ze stymulacją odpowiedzi odpornościowej
przy czym
Prócz tych trzech podstawowych zmiennych w modelu występuje zmienna dodatkowa, która opisuje stopień uszkodzenia organu zaatakowanego przez antygen, ponieważ Marczuk budował swój model z myślą o żółtaczce i stopniu uszkodzenia wątroby w wyniku tej choroby.
Niech
Mamy zatem
gdzie
Wielkość
Ostatecznie model Marczuka humoralnej odpowiedzi odpornościowej opisujemy układem 4. równań różniczkowych z opóźnionym argumentem (rrzoa)
| (13.1) |
W większości przypadków istnieje pewna progowa wielkość
![Przebieg typowej funkcji $\xi(m)$, czyli funkcja tożsamościowo równa 1 na odcinku $[0,m^{*}]$ oraz malejąca liniowo do 0 na $[m^{*},1]$.](wyklady/mbm/wyk13_przykladowe_eps_od_m.hd.png)
13.2. Podstawowe własności układu (13.1)
Przejdziemy teraz do zbadania podstawowych własności układu (13.1). Jest on układem równań z opóźnionym argumentem, zatem jako warunek początkowy należy zadać funkcję określoną na odcinku
Istnienie, jednoznaczność i nieujemność rozwiązań dla nieujemnego warunku początkowego możemy wykazać stosując metodę kroków. Przypomnijmy, że opiera się ona na zasadzie indukcji matematycznej i pozwala zastosować znane techniki dotyczące równań różniczkowych zwyczajnych do analizy rrzoa.
Istnienie lokalnego rozwiązania układu (13.1).
Niech
| (13.2) |
gdzie
Nieujemność lokalnego rozwiązania układu (13.1).
Pokażemy najpierw, że rozwiązanie to jest nieujemne. Pierwsze równanie układu (13.2) zapisujemy w równoważnej postaci całkowej
gdzie w powyższym wzorze
Dla
| (13.3) |
Ponieważ
W ogólnym przypadku, gdy
Zatem
W analogiczny sposób dowodzimy nieujemności zmiennej
Skoro
Zatem
Istnienie rozwiązania układu (13.1) na przedziale 0 , τ
Pokażemy teraz, że z nieujemności rozwiązań wynika ich istnienie na całym przedziale
zatem wzrost
Dla zmiennej
czyli
Z ograniczoności
Globalne istnienie rozwiązań układu (13.1).
Stosując metodę kroków zakładamy, że istnieje nieujemne rozwiązanie
układu (13.1) na odcinku
Punkty krytyczne układu (13.1).
Znajdziemy teraz punkty krytyczne badanego układu. Rozwiązujemy następujący układ równań algebraicznych
| (13.4) |
W ogólnym przypadku, aby rozwiązać układ (13.4), musimy zadać konkretną postać funkcji
Łatwo widać, że układ (13.4) ma rozwiązanie opisujące stan zdrowia, czyli
Niech
Jeśli
Punkt krytyczny
z czego wynika, że jeśli
-
[A.]
-
-
to
| (13.5) |
zatem
Lokalna stabilność punktów krytycznych układu (13.1).
Badanie lokalnej stabilności punktów krytycznych rrzoa nie różni się w początkowej fazie od analogicznej analizy dla rrz. Niech
| (13.6) |
Linearyzujemy układ (13.6) wokół punktu
| (13.7) |
W kolejnym kroku znajdujemy rozwiązania układu (13.7) postaci
| (13.8) |
Układ (13.8) ma nietrywialne rozwiązania, jeśli
| (13.9) |
Twierdzenie o linearyzacji gwarantuje, że jeśli wartości własne, czyli rozwiązania równania (13.8) mają części rzeczywiste ujemne, to rozwiązanie
Zbadamy wartości własne dla poszczególnych punktów krytycznych. Zaczniemy od
czyli wartości własne są równe:
Wniosek 13.1
Jeśli
Widzimy, że stabilność rozwiązania
Badając stabilność rozwiązania
| (13.10) |
gdzie
| (13.11) |
a wyznaczenie współczynników pozostawiamy jako ćwiczenie. Przypomnijmy, że dla
Wiemy, że jeśli rozwiązanie
jest stabilne dla
| (13.12) |
Warunek
Zauważmy, że zmiana stabilności może nastąpić tylko gdy pseudowielomian charakterystyczny (13.10) ma pierwiastki czysto urojone
czyli
Zdefiniujmy funkcję pomocniczą
Jej dodatnie miejsca zerowe determinują potencjalne zmiany stabilności punktu krytycznego
Zmiana stabilności następuje przy takiej krytycznej wartości
Wniosek 13.2
Jeśli spełnione są nierówności (13.12) i istnieje dodatni punkt krytyczny
Przypomnijmy, że z nierówności
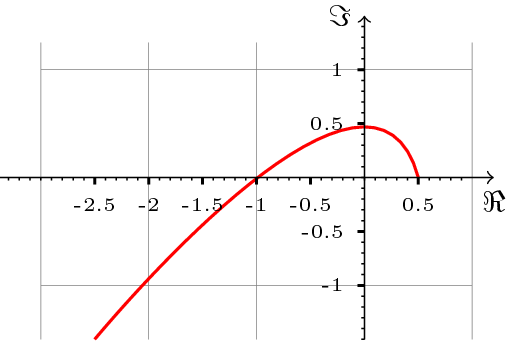
Korzystając z kryterium Michajłowa, można udowodnione następujące twierdzenie dotyczące stabilności punktu
Twierdzenie 13.1
Niech
| (13.13) |
Jeśli dodatkowo spełnione są nierówności (13.12) i istnieje dodatni punkt krytyczny
Twierdzenie 13.1 zadaje warunki wystarczające stabilności punktu
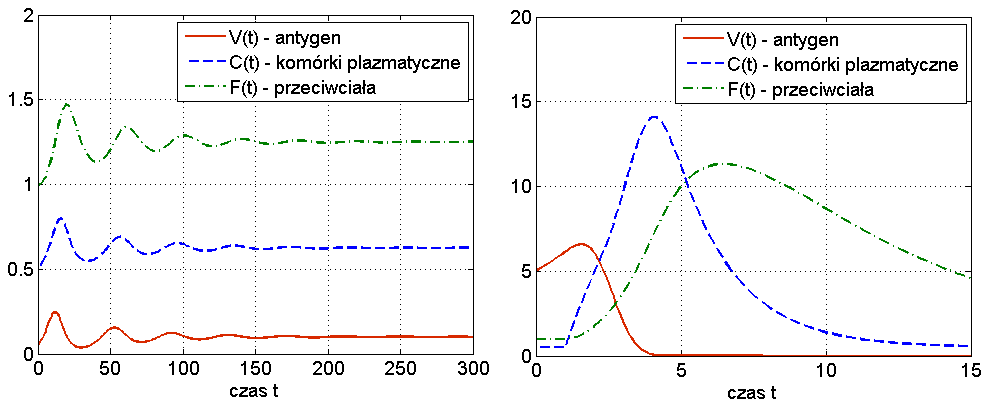
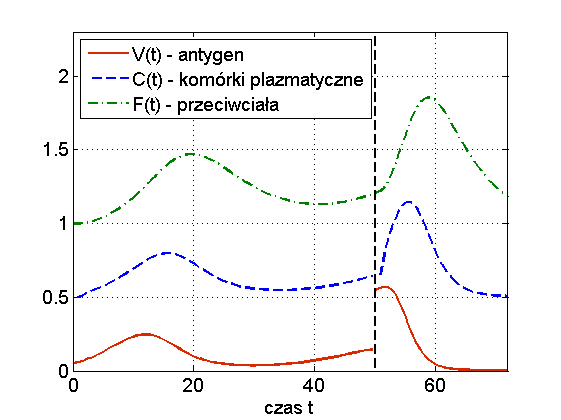
Widzimy więc, że układ (13.1) jest w stanie odzwierciedlić dwa podstawowe przebiegi choroby: zakażenie z wyzdrowieniem oraz chorobę przewlekłą (chroniczną), por. rys. 13.3. Ciekawą własnością symulacji tego modelu jest to, że zwiększając dawkę antygenu możemy wyprowadzić organizm z choroby chronicznej — mówimy o wyzdrowieniu poprzez zaostrzenie choroby, por. rys. 13.4. Trzeba jednak podkreślić, że taki efekt obserwuje się tylko w symulacjach, natomiast wszystkie wyniki analityczne wskazują na globalną stabilność punktu