6. Modele oddziaływań między dwiema populacjami I
Przejdziemy teraz do omówienia zagadnień związanych z modelowaniem ekosystemu, w którym występują dwie populacje. W zależności od oddziaływań między tymi populacjami, rozróżniamy trzy typy ekosystemów
-
układ drapieżnik – ofiara, w którym jeden z gatunków ekosystemu jest pożywieniem dla drugiego;
-
konkurencja, w którym gatunki rywalizują o zasoby środowiska (pożywienie, miejsca lęgowe itp.);
-
symbioza — zjawisko współżycia przynajmniej dwóch gatunków, które wpływają pozytywnie na siebie (mutualizm) lub wyraźne korzyści odnosi jedynie jedna ze stron, nie szkodząc drugiej (komensalizm).
W tym rozdziale zajmiemy się opisem średnich zagęszczeń obu populacji, nie uwzględnimy przestrzennego rozmieszczenia osobników w ekosystemie, co oczywiście kryje dodatkowe założenie, że osobniki są rozmieszczone jednorodnie i jest ich dostatecznie dużo, aby można było mówić o średnim zagęszczeniu.
6.1. Model Lotki – Volterry
Wprowadzimy najpierw pierwszy historycznie model opisujący oddziaływania dwóch populacji w ekosystemie. Dotyczy on układu drapieżnik – ofiara i został zaproponowany równolegle jako model populacyjny przez Vito Volterrę [17] oraz jako model łańcucha reakcji biochemicznych przez Alfreda Lotkę [10]. Volterra zaproponował ten model w celu wyjaśnienia pewnego, zdawałoby się paradoksalnego zjawiska, które zostało zaobserwowane po I wojnie światowej. Po ustaniu działań wojennych, kiedy ludzie na powrót zajęli się uprawianiem swoich zawodów, rybacy odkryli, że populacja ryb drapieżnych w Adriatyku zwiększyła się. Uznano to za paradoks, gdyż zdawałoby się, że wszystkie gatunki powinny ucierpieć w wyniku działań wojennych. Volterra na bazie swojego modelu wykazał, że wzrost liczebności drapieżników był całkiem naturalny, ponieważ w czasie wojny zaprzestano połowów i dzięki temu populacja drapieżników mogła wrócić do stanu naturalnego. Co więcej, model ten odzwierciedla znane w ekologii prawo zachowania średnich, które mówi, że w naturalnych siedliskach zmiany liczebności populacji w czasie zachodzą tak, że zachowana zostaje liczebność średnia.
Zaczniemy od przedstawienia modelu heurystycznego, na bazie którego wyprowadzamy równania modelu Lotki – Volterry. Zakładamy, że w ekosystemie występują dwa gatunki
Niech
| (6.1) |
gdzie dla uproszczenia zapisu pomijamy zmienną niezależną
-
-
-
współczynnik
Przejdziemy teraz do analizy zachowania rozwiązań układu (6.1). Ponieważ układ (6.1) jest autonomiczny, więc bez straty ogólności przyjmujemy, że
Stwierdzenie 6.1
Rozwiązania układu (6.1) dla nieujemnego warunku początkowego
Ponieważ prawa strona układu (6.1) jest klasy
Pozostaje jeszcze udowodnić istnienie rozwiązań dla wszystkich
Po pomnożeniu powyższej nierówności przez
czyli
Zauważmy, że gładkość prawej strony i liniowe oszacowanie pochodnej zawsze gwarantuje przedłużalność rozwiązań.
Przystąpimy teraz do analizy asymptotyki rozwiązań układu (6.1). Wyznaczymy najpierw stany stacjonarne
zatem albo
Dla zerowego stanu stacjonarnego otrzymujemy
widzimy więc, że jest to punkt siodłowy, gdyż wartości własne
czyli wartości własne macierzy Jacobiego
Spróbujmy naszkicować portret fazowy. Zaczniemy od wyznaczenia izoklin zerowych, czyli krzywych, na których
oraz
Izokliny zerowe dzielą przestrzeń fazową
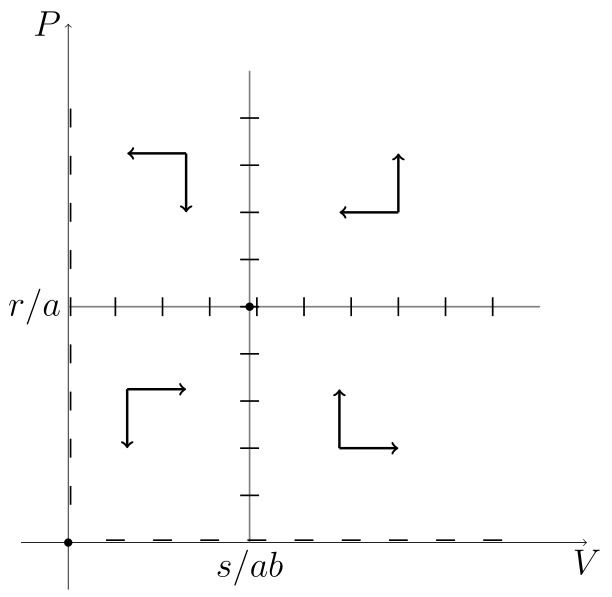
Można pokazać, że wszystkie rozwiązania są okresowe, co wynika z własności całki pierwszej [13] układu (6.1). Znalezienie całki pierwszej i zbadanie jej własności pozostawiamy jako ćwiczenie.
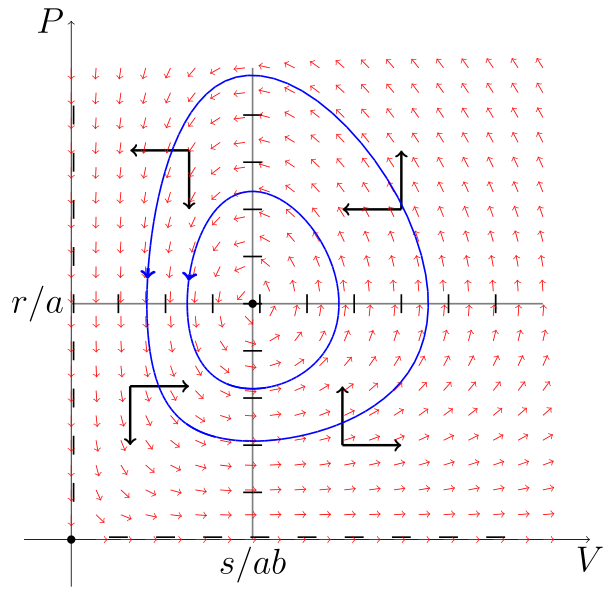
Na koniec zauważmy, że wszystkie rozwiązania oscylują wokół stanu stacjonarnego
a ponieważ rozwiązanie jest okresowe, to
Analogicznie pokazujemy, że
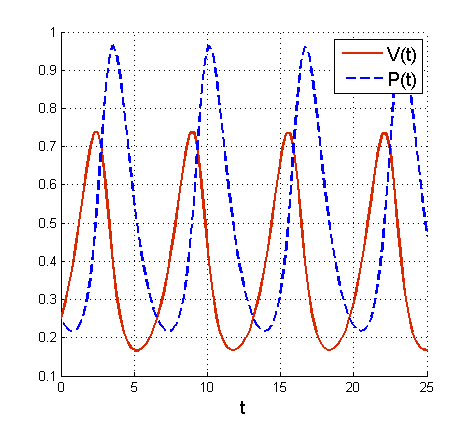
Na tej podstawie wyjaśnimy paradoks związany z populacją ryb drapieżnych w Adriatyku. Zauważmy, że jeśli odławiamy ryby, to zakładając jednakową intensywność odłowu dla obu populacji
| (6.2) |
gdzie
zatem odławianie powoduje zmniejszenie średniej liczebności populacji drapieżników i zwiększenie średniej liczebności populacji ofiar. W związku z tym w czasie gdy nie było połowów, populacje wróciły do stanu naturalnego, więc liczebność drapieżników wzrosła.
6.2. Konstruktywna krytyka modelu Lotki – Volterry
Chociaż model Lotki – Volterry odzwierciedla obserwowane w naturze oscylacje w układach drapieżnik – ofiara, a także w sposób analityczny dowodzi ekologicznego prawa zachowania średnich w tych układach, to jednak nie jest pozbawiony wad. Przede wszystkim z matematycznego punktu widzenia jego główną wadę stanowi brak stabilności strukturalnej. Pojęcie stabilności strukturalnej wykracza poza ramy tego wykładu — w skrócie stabilność strukturalna oznacza, że niewielka zmiana prawej strony układu nie prowadzi do radykalnych zmian w dynamice rozwiązań. Widzimy, że w przypadku układu (6.1) nawet bardzo mała zmiana prawej strony może skutkować zmianą typu stanu stacjonarnego — stan typu centrum bardzo łatwo zaburzyć. Dodatkowo model ma też pewną własność, która podlega krytyce z ekologicznego punktu widzenia. Rozwiązania układu (6.1) oscylują w taki sposób, że wzrost populacji drapieżników poprzedza wzrost populacji ofiar, natomiast w większości rzeczywistych układów drapieżnik – ofiara takie oscylacje są przesunięte w stosunku do tych rozwiązań; najpierw obserwujemy wzrost populacji ofiar, a potem następujący po nim wzrost populacji drapieżników. Należy też zauważyć, że w modelu heurystycznym pominięto wiele istotnych dla układów drapieżnik – ofiara czynników. W związku z tym model Lotki – Volterry był na wiele różnych sposobów modyfikowany i poniżej omówimy pokrótce kilka z tych modyfikacji.


