Zagadnienia
1. Stochastyczne modele ekspresji genów
1.1. Krótki wstęp biologiczny
Jednym z podstawowych procesów biochemicznych zachodzących w komórkach żywych organizmów jest produkcja różnych białek. Jest to bardzo skomplikowany wieloetapowy proces. Każda cząsteczka białka składa się z połączonych w liniowy sposób odpowiednich aminokwasów. Opiszemy teraz w niezwykle uproszczony sposób jak dochodzi do połączenia aminokwasów w cząsteczkę białka (por. Rys. 1.1). Informacja genetyczna
umieszczona jest na nici kwasu DNA o strukturze podwójnej helisy. Wzdłuż nici DNA rozłożone są komplementarne zasady,
odpowiednio ATCG. Kod genetyczny jest kodem trójkowym, trzy następujące po sobie zasady kodują odpowiedni aminokwas.
Zauważmy, że mamy
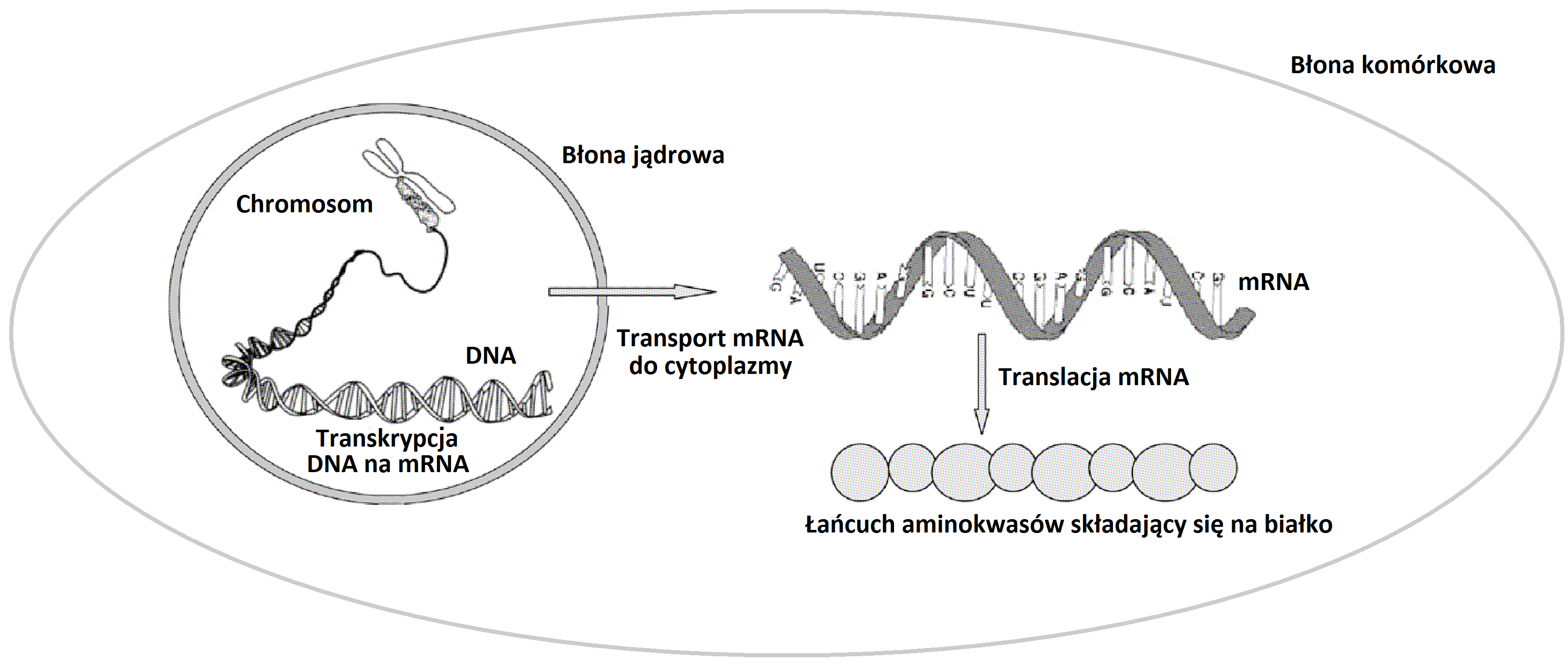
Uzyskane białko może pełnić bardzo różne funkcje: budulcowe, transportowe, regulatorowe. Regulacja białkowa może polegać na wzmacnianiu (aktywacji) lub osłabianiu (represji) produkcji innych białek lub też samego siebie (autoregulacja). Ostatnio odkryto, że podobne funkcje regulacyjne mogą również pełnić cząsteczki RNA.
1.2. Matematyczny model ekspresji genu
1.2.1. Opis deterministyczny na poziomie makro
Stan komórki jest opisany przez zależne od czasu koncentracje mRNA
| (1.1) |
gdzie
| (1.2) |
1.2.2. Opis stochastyczny na poziomie mikro
W wielu komórkach, zwłaszcza prokariotycznych, liczba cząsteczek mRNA i białek może być niewielka
i w związku z tym mówienie o koncentracji traci sens. Mamy do czynienia ze skończonym układem czasami nawet kilku
cząsteczek danego białka i w związku z tym bardzo duża rolę odgrywają fluktuacje stochastyczne związane
z losowymi czasami zajścia odpowiednich reakcji biochemicznych. Na poziomie mikroskopowym stan komórki opisujemy
zależną od czasu liczbą cząsteczek mRNA
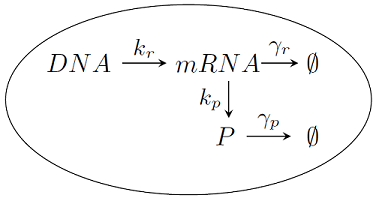
Reakcje te opisywać będziemy przy pomocy procesu urodzin i śmierci (patrz Dodatek 1).
Przyjmujemy następujące prawdopodobieństwa zajścia reakcji w odcinku czasowym
-
transkrypcji, czyli przejście
-
translacji, czyli przejście
-
degradacji mRNA, czyli przejście
-
degradacji białka, czyli przejście
Naszym celem jest znalezienie wyrażenia na wariancje,
Ćwiczenie 1.1
Skonstruuj nieskończony układ równań różniczkowych zwyczajnych dla
Zastosuj procedurę z Dodatku!
Definiujemy funkcję tworzącą dla prawdopodobieństw
Ćwiczenie 1.2
Skonstruuj równanie różniczkowe cząstkowe dla
Zastosuj procedurę z Dodatku 2.
Oznaczmy przez
Ćwiczenie 1.3
Zróżniczkuj powyższe równania ze względu na
Zauważmy, że powyższy układ równań różniczkowych jest zamknięty, w równaniach dla danego momentu nie występują momenty wyższych rzędów - macierz układu równań jest trójkątna. Możemy więc po kolei rozwiązywać równania różniczkowe (porównaj (?)) i dostać wyrażenia na ewolucję czasową momentów. Zauważmy, że dwa pierwsze równania są takie same jak w układzie równań różniczkowych na koncentracje mRNA i białka w modelu deterministycznym (1.1).
Ćwiczenie 1.4
Znajdź wyrażenie na wariancję liczby cząsteczek białka w stanie stacjonarnym.
1.3. Proces Poissona
Proces Poissona to rodzina zmiennych losowych
Założenia
-
Prawdopodobieństwo wystąpienia zmiany (urodzenia się jednej dodatkowej cząsteczki lub ogólnie wystąpienie pewnego zdarzenia) w odcinku czasowym
-
Prawdopodobieństwo wystąpienia więcej niż jednej zmiany w odcinku czasowym
Zauważmy, że z powyższych założeń wynikają następujące własności:
-
Prawdopodobieństwo wystąpienia zdarzeń w rozłącznych odcinkach czasowych jest równe iloczynowi odpowiednich prawdopodobieństw (niezależność);
-
Prawdopodobieństwo wystąpienia zdarzenia nie zależy od czasu
Oznaczmy przez
Naszym celem jest znalezienie wzoru na
Ćwiczenie 1.5
Skonstruuj równanie różniczkowe zwyczajne na
Napisz wyrażenie na prawdopodobieństwo całkowite.
Wartość zmiennej losowej
Możemy napisać wyrażenie na prawdopodobieństwo całkowite:
Przenosimy
| (1.3) |
z warunkiem początkowym
Rozwiązujemy równanie różniczkowe na
| (1.4) |
Rozwiązanie równania dla
Ćwiczenie 1.6
Sprawdzić, że
jest rozwiązaniem układu (1.3),
to znaczy, że
1.4. Proces urodzin i śmierci
Tak jak i w procesie Poissona
Zakładamy, że
-
Prawdopodobieństwo urodzenia się cząsteczki w odcinku czasowym
-
Prawdopodobieństwo śmierci cząsteczki w odcinku czasowym
Przyjmiemy teraz, że
| (1.5) |
Nie będziemy rozwiązywać tego układu równań różniczkowych. Interesować natomiast nas będzie stan stacjonarny,
1.4.1. Błądzenie przypadkowe na kracie jednowymiarowej z czasem ciągłym
Jest to proces urodzin i śmierci, dla którego
Układ (1.5) możemy wtedy przepisać jako
| (1.6) |
z warunkiem początkowym
Różniczkując funkcję tworzącą ze względu na
gdzie
warunek początkowy
Rozwiązaniem powyższego równania różniczkowego jest
| (1.7) |
Ćwiczenie 1.7
Znajdź wartość oczekiwaną oraz wariancję
Ćwiczenie 1.8
Znajdź


